Show me your genes and I will tell you who you are
Dr. David Heaf
Originally published in New View, 4th Quarter, Autumn 2000, pp
7-12
"Monday 26th June 2000 will be remembered as the day
when humankind learned, in a sense, what it is to be human."1 You might be
forgiven for thinking something momentous has happened. You may even remember for the rest
of your life what you were doing when the news reached you. Certainly some important
people thought something important had happened. Bill Clinton said, "This is the most
wondrous map ever produced by mankind." Tony Blair called it, "a breakthrough
that opens the way for massive advancement in the treatment of cancer and hereditary
diseases, and that is only the beginning."
So what in fact had happened? The leaders of the publicly
sponsored Human Genome Project (HGP) and Craig Venter who leads the parallel private
initiative of the company Celera Genomics chose that day to announce that they had
completed the first working draft of the complete set of human genetic information, the
human genome (see Box 1 for other basic terms). This feat has been likened to the 1953
conquest of Everest, but when what the HGP has accomplished is examined, a more apt
comparison would be setting up base camp for the final ascent. 85% of the genome has had
its base sequence worked out roughly and 24% has been sequenced to 99.99% accuracy, a
standard that needs the whole sequencing process to be repeated about 10 times (see Box 2
on sequencing). Furthermore, with some 3.1 billion base pairs in the genome even when the
whole genome has been sequenced to that accuracy, there could remain some 310,000 errors,
and these in only a 'generic' genome of the few who contributed their DNA to the sample.
There remain too the genetic differences between human individuals. These are important if
Tony Blair's prediction for disease treatment is to be realised. These differences, most
of which are accounted for by 'single nucleotide polymorphisms' (SNPs),2 are
also receiving intensive research and with the extraordinary acceleration of the project
through mechanisation combined with computing power, sequencing the genomes of individuals
may be only a few years off.3 The remaining 15% of the genome which has not yet
been sequenced is likely to prove very resistant to sequencing by present methods for
technical reasons arising from the repetitive nature of the sequences it contains.4
Even imagining that the 26th June was the announcement of the accurate
sequence, nobody knows what 97% of it does and only 38,000 genes are as yet known, less
than a third of what may yet be discovered. So to return to our Everest metaphor, no
summit flag has yet been planted.
Why then all the song and dance now? One likely factor is the
last-minute rapprochement in Spring 2000 between the private and public efforts which
until then appear to have been competing against each other. "An unseemly feud would
have marred their effort to present the 'mapping' of the human genome as a noble and
high-minded endeavour."5 Another likely factor is that most biotechnology
research ventures need to keep up the hopes of investors – public and private –
in order to keep the cash coming in. The final decision to sequence the whole genome was
taken in 1990. In 1991 in the USA alone $130 million was spent on it. In 1998 the
expenditure was $250 million. And there are now teams in some 18 other countries involved
in the effort. Britain's Sanger Centre at Cambridge, funded to the level of £210m by the
Wellcome Trust, contributed about a third of the sequence data obtained so far.6
Even when the whole sequence is known, which could be many years away, will we have the
'essence of mankind'?7
| Box
1. Chromosomes, genes and DNA
Basics: Figure 1 shows an artist's impression
of an electron microscope picture of a typical representative of one of the 100 trillion
or so cells in the human body. Coming from one of the chromosomes of the 23 pairs in the
cell's nucleus is a purely diagrammatic representation of a chain of DNA (Deoxyribose
Nucleic Acid). Each of the two strands in the DNA double-helix chain is made of a backbone
of repeating sugar and phosphate molecules (shown as a helical ribbon in Fig. 1).
Chemically, DNA is thus a polymer. To each sugar molecule is attached one of four
molecules called bases, designated by the letters, C, A G and T, the first letters of
their chemical names cytosine, adenine, guanine, and thymine. The group comprising
phosphate, sugar and attached base is called a nucleotide. Bases on opposite complementary
strands pair up, T always with A, C always with G. The weak attraction or bonding between
the two bases of the pair help to hold the two strands of the helix together. It is this
pairing and bonding of the bases combined with their linear distribution along the DNA
strand which makes it possible to determine the sequence of bases in DNA. There are some
3.1 billion base pairs in human DNA. The chromosomes vary in size having 50-250 million
base pairs each.
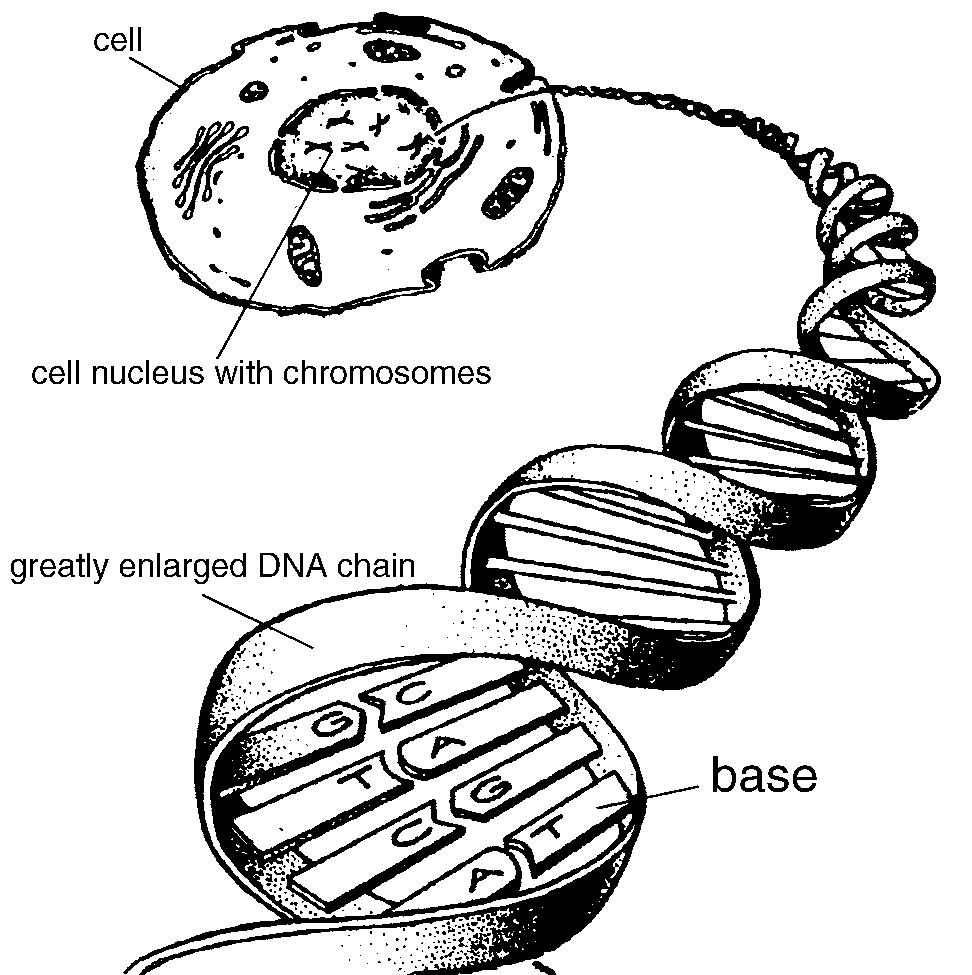
Figure 1: Artists impression of a cross section of a cell with
chromosomes and DNA
The gene: a fuzzy concept. A classical
geneticist sees a gene as a unit of heredity transmitting definite characteristics (e.g.
eye colour) from parent to offspring according to Mendel's laws of inheritance. To a
molecular biologist it is a stretch of bases on a DNA strand that codes for a unit of
function in the cell. An evolutionary biologist regards a gene as a cell component robust
enough to serve as the basis for evolution. And the biochemist's gene is a piece of DNA
that the cell needs in order to make a protein, which could be either an enzyme needed to
facilitate a chemical reaction or a protein which is part of the structure of cells and
tissues of the body. Proteins are built up of chains of some 20 different amino acids,
each coded for by its own triplet of adjacent DNA bases from among the foursome C, A, G
and T.
The gene concept gets even fuzzier when the following properties are
considered: Genes can split into several parts along the DNA chain or have base sequences
which overlap with other genes. They often have no defined ends because of their
associated flanking and control sequences. The fluidity of the genome and so called
'jumping genes' means that genes often have no permanent location in chromosomes or DNA.
The totality of the genes making up the human genome is not yet known, but is likely to be
between 40,000 and 140,000. Furthermore, 97% of the DNA is thought not to code for
proteins and is sometimes referred to as 'junk DNA'. It probably plays an important part
in the dynamic and structural organisation of the DNA such as controlling the expression
of genes.
There is a 98.5% match between human and chimpanzee DNA and a 99.9%
match between the DNA of different human individuals. According to the molecular
geneticists then, individual genetic differences, including maleness and femaleness are
therefore largely attributable to the remaining 0.1% of the DNA. |
Box 2. Mapping and Sequencing
the Human Genome
Mapping: One of the earliest maps of the human genome came from
studying how characteristics are passed on in families. If two (or more) characteristics
tend to crop up together, we might suppose that they were coded for by genes on the same
chromosome. However, even if two characteristics are coded for on the same chromosome,
occasionally they will occur separately because of the occasional jumbling up of the
chromosomes (meiotic crossing over) that happens when sperm cells and egg cells are
formed. If two characteristics that are on the same chromosome get separated frequently,
then we can conclude that they are far away from each other on the chromosome. If they
hardly ever get separated, then the two characteristics must be very close together on the
chromosome. By looking at how characteristics are inherited in patterns, we can draw up a
vague map of where the genes are on the chromosomes. The disadvantage of this approach is
that humans take a long time to grow up and have children, so it is only usually possible
to look at three or four generations of a single family at a time. Also, there are often
not enough children in each family to work out the gene positions accurately.
Whilst the chromosome maps from this classical genetics approach provide useful
orienteering information, to obtain the actual base sequences of the DNA in the genes of
the chromosome, molecular biological techniques are necessary.1 One such technique
involves working backwards from the protein coded for by a gene. There is a correspondence
between the genetic code of bases in DNA and the amino acids, the building blocks of
protein. From the amino acid structure of a protein a short strand of DNA can be
synthesised, called a probe, which when mixed with the chromosomes or DNA containing the
gene coding for the protein will bind firmly to it. If the probe has a marker attached to
it which can be seen by microscopy, the position the marker shows on the chromosome or DNA
indicates the position of the gene.
Sequencing: Because of the high degree of similarity of DNA from
different people, DNA for working out its sequence of bases was obtained from the sperm
and blood of only a dozen or so anonymous volunteers and extracted into a watery solution
with added chemicals to protect it.2 Using enzymes it was 'cut up' into lengths of about
150,000 bases to make a living 'library' of human DNA stored in artificial chromosomes in
bacteria (BACs). By bacterial cloning techniques the BACs can be multiplied, kept pure,
sampled when needed and the human DNA recovered. As the human DNA in the BACs is still too
long for the sequencing step it is cut into smaller fragments of 400-700 bases and this is
done so that the sequences of the fragments overlap with one another. To obtain sufficient
material for the next step each of the fragments is copied many times using the polymerase
chain reaction (PCR).
Any length of DNA can be copied, provided it has a start signal at one end and a
stop signal at the other end. The DNA is added to a test tube full of DNA ingredients,
such as the four bases (A, C, G and T), sugars and phosphate. DNA polymerase, the enzyme
that catalyses the formation of DNA, is also present. The mixture is heated, so that the
weak bonds between the bases in the two strands of the DNA (see Fig. 1) are broken. Once
these strands are separated, the mixture is cooled and each strand can act as a template
for DNA polymerase, so complementary sequences are soon generated for each of the original
strands. Then the mixture is heated again, so that the bonds are broken and the strands
separate. This time, there are four strands of DNA to act as templates once the mixture is
cooled. The cycle of heating followed by cooling is repeated many times, and the amount of
DNA is doubled with each cycle.
To sequence a length of DNA, a copy of the DNA to be sequenced is
added to four test tubes. Within each of these test tubes are all the ingredients to make
new DNA (see previous para.). There is, however, one important difference between the
contents of these test tubes, and the usual ingredients for making DNA. The difference is
that in each of the test tubes, one of the bases – A, C, G or T – has been
replaced with a modified version of the base which when inserted prevents the DNA strand
from growing further. In the test tube where A has been replaced with the modified version
of A, there will be DNA fragments of different lengths, each one terminating in one of the
modified bases. Wherever there is an A in the original sequence, some of the growing
chains of DNA in the A test tube will have been stopped at that position.
To identify the fragments of DNA that have been produced, the DNA pieces are
separated in a gel through which the movement of DNA fragments is partially impeded
because it is made up of long, polymer molecules. An electrical field is applied across
the gel, so that one end of the gel is negatively charged and the other end is positively
charged. The pieces of DNA are added to the gel at the negatively charged end. Because DNA
is slightly negatively charged, the bits of DNA begin to move through the gel, away from
the negatively charged end and towards the positively charged end. The speed with which
the DNA moves depends on its size – the smaller pieces of DNA are able to move more
quickly through the gel. After a while, the pieces of DNA are spread out across the gel.
The smallest ones have nearly reached the positively charged end while the largest pieces
have hardly moved from their starting position.
Modern sequencing machines have the gel in capillary tubes. The reactions in our
four test tubes can be combined and added to one capillary. In which case the emerging DNA
chains, terminating in modified A, C, G or T, are detected electronically from the light
they emit when mixed with fluorescent dyes specific for the four different bases and
illuminated by a laser. The results are fed into a computer. Some labs still use
multi-lane gels run between two glass plates. Here detection can be by radioactive tagging
followed by radiography of the gel. In this case the contents of the four tubes are kept
separate and placed in the gel's electric field in four separate lanes.
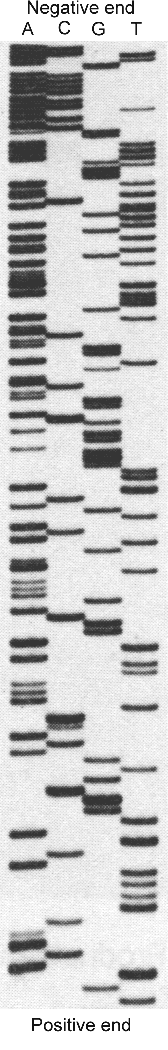
Figure 2: Radiograph
of an electrophoresis gel of fragments of DNA showing the four lanes corresponding to the
bases A, C, G & T. Each dark horizontal band on the radiograph is produced by the
radioactive tag in a short strand of DNA, the longest strands at the top and the shortest
at the bottom. The direction of flow was from top to bottom. The sequence, 'read' from the
bottom, is TGTACAACTTTTA etc.
The base sequence emerging from the capillary or 'read' from the gel (see Fig. 2 above)
is the sequence of just one of the millions of DNA fragments created at the beginning. The
fragment sequences are fed into powerful computers programmed to do a huge linear 'jig-saw
puzzle' by working out what the sequence of the DNA chain was before it was cut into
fragments. The following diagram illustrates with three fragments of DNA how use is made
of the overlaps in the sequences of the fragments:
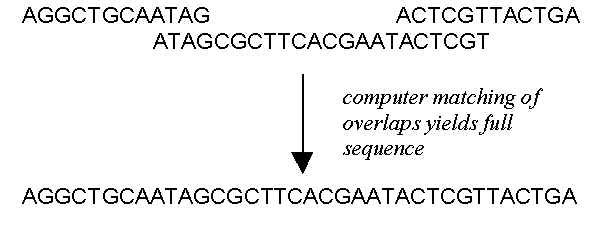
Notes
1. The Sanger Centre web site: www.sanger.ac.uk/HGP/draft2000/how.shtml
2. www.sanger.ac.uk/HGP/draft2000/who.shtml and www.ornl.gov/hgmis/faq/seqfacts.html#whose
|
DNA thinking
It was recognised at the outset of the HGP that once it had
reached its goal it would have ethical, legal and social implications (ELSI). The ELSI
program ran in parallel from the beginning, drawing on some 3-5% of the overall funding of
the project. Already by 1991 the key concerns were being discussed in workshops: eugenics,
racism, genetic discrimination, genetic determinism, reductionism, control of genetic
information, screening, counselling, impact on procreative liberty, patent rights, gene
therapy and enhancement.8 As this ground has already been dug over so much in
the press, both lay and technical, we shall not visit it again here. For an excellent
insight into the key issues the reader is referred to Bryan Appleyard's Brave new
Worlds: Genetics and the Human Experience.9 As we have seen, the HGP is far
from completion and has thus not had time to do any damage. We can only speculate at this
stage what some of the consequences might be, good or evil. But we can look at it for what
it is in itself, intrinsically, in principle. We can examine the world views behind it and
its proposed applications. In doing so we come to the difficulties science has with
understanding what being human and becoming ill mean, which will be the main focus of the
remainder of this article.
The biological sciences have laboured for more than two centuries
under a methodological problem in their underlying thinking. They have tended to assume
that the mechanistic thinking which works well in physics can be transferred with a few
modifications to biology. And biology has now become the study of the behaviour of
molecules. It has long since given up any aspiration to cognitive faculties appropriate
for the understanding of life. We can distinguish three different cognitive approaches
which belong to different realms of science. In the science of physics we can find all the
circumstances which are necessary to bring about a given phenomenon and by thinking about
the connections arrive at a law. We can seek out primal phenomena in which the law becomes
visible. Such is the approach to inorganic nature. But in organic nature, the realm of
biology, to reach a comparable level of certainty we need to be able to apprehend the type
at work in the organism concerned and follow it through in all its ramifications in the
forms and functions under study. This is not to say that organisms are not also subject to
the laws of physics. When we come to the human being, we rise to a new level in the
scientific process. Of course, as with any animal, there is much about human beings which
is purely representative of the type, but the science of the human being needs to take
into consideration an additional factor – the individuality. Here individuality takes
on the role that type does for animals. Here we have briefly outlined a Theory of
Knowledge Implicit in Goethe's World Conception as set out by Rudolf Steiner in 1886.10
Indeed, 'Down to the smallest particle of his substance, man in his form and configuration
is a product of the organisation of the Ego'.11 Here is meant the essential
self, that principle of individuality in each person. So just as the animal type stamps
itself on the base sequences of its DNA, so too the human individuality, working as the
incarnate ego through forces which form the physical body, can to a greater or lesser
degree stamp itself on the 'smallest particles of his substance', the composition of his
DNA. Therefore, when we have sequenced the whole of the generic human being, we will still
be a very long way from knowing the 'essence of mankind' or 'what makes us human', as
James Watson the co-discoverer of the DNA double-helix described the molecule.12
That essence, the human is the individual. And in all our considerations of what
should be done with the information pouring hour by hour into the databases of the HGP at
the very least we should not overlook that fact.
In thinking 'what it is to be human' is coded by the DNA, even if
this idea is meant 'in a sense', we divert attention away from the reality of what it is
to be human. We fall into the ongoing error in biology of collecting data, applying
statistics and reaching generalisations within degrees of probability, albeit the 99.99%
probability of the HGP's 'gold standard', which even if we were studying an animal, would
leave us blind to the type manifesting in our sample. We become dependent on the inductive
method. We collect observations of a sufficient number of instances and arrive at a
generalisation about them which is always only provisional because a new instance may turn
up which forces us to modify our generalisation. But once the type has been
grasped, the result is never provisional. This is best illustrated by Goethe's correct
prediction and discovery of the human premaxilla (intermaxillary bone) despite the
scientific opinion of his time which held that the absence of the bone distinguished man
from animal. He correctly realised from the inner lawfulness of the type that the human
form too must manifest the bone.
The HGP takes the study of anatomy to its ultimate extreme. As
with most anatomical research, what was once living has to be killed and fixed – what
once in-formed it is driven out. This shows vividly in Rembrandt's painting The Anatomy
Lesson of Dr. Tulp (Figure 2. below) in which the various gazes into the middle
distance seem to convey an atmosphere of wonderment for the whereabouts of the departed
soul. In extracting and preserving the DNA, the sequence of the human genome at best
represents a snapshot in the temporal stream that characterises every living process. It
gives us human molecular anatomy of the 21st century and thus cannot be seen as
finished for all time. After all, even the summit of Everest is not a fixed point, neither
in space nor above sea level.
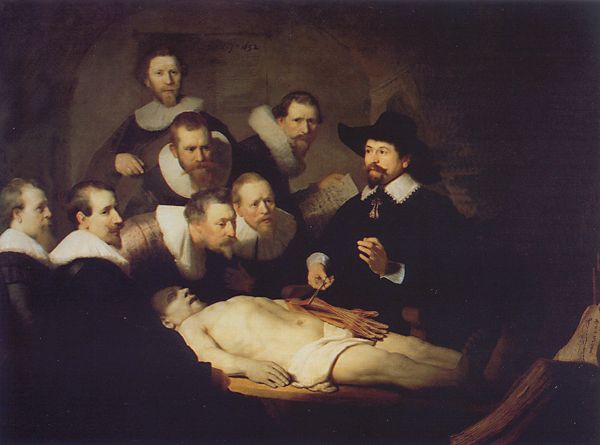
Figure 2: The Anatomy Lesson of Dr. Tulp by Rembrandt
Understanding that the human individuality stamps itself on its
manifest form right down to the level of the DNA admittedly involves a radical departure
from current western thinking about the human being. It involves looking at a broader time
span than a single life. We are used to thinking of heredity as involving something being
passed on to us by our parents. It is something done to us, so to speak. But Rudolf
Steiner describes from spiritual scientific observation how the incarnating individual
works down through the stream of heredity many generations in advance of the one in which
he will manifest on earth, preparing the qualities he is ultimately to inherit. In a
sense, he inculcates these qualities into his ancestors – albeit in concert with
other beings, including future siblings, who are active in his hereditary stream –
until a physical body can be born to which he feels drawn.13 Dominic Lawson,
former editor of The Spectator had a sense for the deep connection of individuality
and bodily form when he wrote:
"Two emotions coursed through me as the consultant gave me a
guided tour of the stigmata of Down's Syndrome. The first was anger. While I understood
that the doctor was only doing his professional duty – to explain as clearly and as
quickly as possible the condition of his patient – I wanted to shout out, " This
is my daughter you are prodding, not some random strip of flesh." The second emotion
was love...I felt an intense, almost physically painful love for this third
daughter....the Down's baby is as much the product of his or her parent's genes as any
other child... There is no possible alternative Domenica Lawson without Down's Syndrome.
That is her identity, her very essence, along with all the other genes she has inherited
from us."14 And yet no matter how individual it is, the DNA cannot be the
origin of the individuality. This runs counter to the view amongst certain biologists that
higher order attributes of an organism are an 'emergent' property of the lower order
attributes. Put simply, properties of molecules are supposed to emerge from the
association of its individual atoms, properties of organs from collections of molecules,
consciousness from interaction of organs and so on. It is reductionism in new clothes. In
contrast to this, ideas of causation originating from Aristotle and, later in the
Middle-Ages, Thomas Aquinas, are used in Catholic ideology to argue that the human soul is
present in the embryo right from the moment of conception.15 Their point being
that cells in the embryo cannot 'cause' the higher level of order of the soul. The soul
provides the ordering influence from the start, right down to the level of the cells.
Personhood starts at conception.16 Rudolf Steiner too describes how the
ordering element, the form of the human body – what he calls the 'spirit germ' enters
at conception. This cosmic origin of form thus transcends heredity. But in contrast to
Catholic ideology, Steiner points out that the spirit soul elements delay their entry into
the embryo until the 17th - 20th day.17 Incidentally it
is worth noting here that in the UK, experimentation on the human embryo is permitted only
up to the 14th day of development when a visible change takes place in the
embryo. The primitive streak forms, evidencing the rudiments of the nervous system and
marking the beginning of a new phase of development.
The DNA-thinker has been seduced by the admittedly huge
informational complexity of the DNA, albeit with its simple code based on A, C, G or T.
Molecular biological dogma long held that information flow was from DNA to protein to
organic form. The DNA was supposedly the blueprint. More and more phenomena are being
discovered that erode this simplistic picture. Not only does control information flow in
the reverse direction – that is from cell periphery to the DNA –, but also
processes totally outside the DNA, so called epigenetic processes play an important
part in the stream of heredity.18 With the help of enzymes, the organism
manages its DNA to suit its purposes. The DNA comes into existence, is repaired, copied
and maintained by the organism. Like all substances in the organism, it is in a state of
constant flux and turnover, something which the fixed models such as in Fig. 1 conceal
from our imagination. And no single molecule, despite its size or complexity, deserves to
be given pride of place in this organismic view. The organism is primal. But such a
thought sorely challenges the DNA-thinker. Where then is the organism? What
material object represents it? And the answer is of course that it is a nexus of relations
between perceptible entities and events which can be grasped only in the ideal realm of
human cognition, and yet is no less real for that.
A further problem with DNA-thinking is the idea that what happens
now is programmed by something which happened in the past. 'Our fate is in our genes'
thinks Watson.19 This shows the kind of concept of causality which is
appropriate only to inorganic nature. The bullet moves impelled by a previous
explosion. But life is not like that. It no doubt takes up what is there, but its fluidity
transcends such narrow ideas of causality. The river of life flows round obstacles.
Appleyard in the book already mentioned touchingly describes this from his own experience
of how his niece lived her shortened life to the full despite the fate prepared for her in
her genes. And such destiny events are in their turn an opportunity for transformation for
all those caring for the sufferer.
So in contrast to whatever 'corrosive'20 powers have
been dreamed by reductionists into Deoxyribose Nucleic Acid to dissolve away any
idea of higher order, of a true human essence or entelechy, we can see that the DNA is of
secondary importance in forming the human being. It is no less necessary for that, as are
formative factors such as nourishment, warmth and the social context. Indeed, disturbances
in the DNA are often associated with dramatic changes in the form and function of the
human being, an aspect we shall come to later.
Genomic medicine
Discussing illness in the context of genes soon leads us into the
nature versus nurture debate, or in this context to trying to establish whether an illness
is genetic or environmental. But a science of the human being, anthroposophy, finds
neither of these poles of presumed explanation or causation adequate. It follows from the
foregoing that a third, most important, factor is involved, namely the incarnating
individuality in relation to which the admittedly indispensable conditions of genes and
environment are subordinate. Man is a master of his heredity, not it's slave.21
Anthroposophical medicine recognises not only the physical body and ego, but also the
subtler 'bodies' working in between them into the physical. Therefore in anthroposophical
medicine, the origins of illness are sought in how harmoniously or otherwise the ego
mediates its use of the body through these subtle bodies.22 Too strong or weak
in-forming by the ego or a predominance of either of the intermediate bodies can leave its
mark on the physical and is manifested as illness. When 'mere heredity prevails', it is a
sign of illness – the ego is shirking its task.23
Neither does an anthroposophical approach ignore the importance of
the genes. Indeed, we have already seen how the genes are an essential part in an
evolutionary-karmic process and are not just an accident that happens at conception.
However, I believe that placing so much hope for the future of medicine on knowing the
human or the individual's genome is a big mistake. Tony Blair hopes that it will advance
the treatment of cancer. But a recent study of medical records of 44,788 pairs of twins
born in Sweden, Denmark and Finland since 1870 concluded that overall, if one identical
twin had cancer, his or her sibling had a less than 10% chance of contracting the same
disease.24 Clearly, this statistic alone shows that knowing genomes is not
going to solve the cancer problem for most people. Incidentally, the authors concluded
from this finding that environmental factors are of overwhelming importance in the
aetiology of cancer, but in the light of comments in the previous paragraph that would be
too simple a conclusion. Taking a broader perspective, we can say that cancer occurs,
where the formative influences of the higher members of the human being, are no longer
able to regulate the aspect of the body responsible for effecting the living vegetative
processes (cf. note 17). This manifests as unformed prolific growth in a tissue. And this
embraces the fact that environmental factors such as chemicals and electromagnetic
radiation (e.g. microwaves) can oppose the working of normal bodily formative processes,
leading to mutations in the genes. It is worth noting in this context that the HGP in the
USA was initiated by the Department of Energy which has continued to contribute a major
share of the funding. The US DoE's interest is in wanting fundamental genetic information
about the human being for research on how environmental radioactivity, for instance from
the nuclear power programme, might affect people via their DNA. We can also question how
'environmental' environmental factors really are. Of all living beings, man has the
greatest ability to create his own environment. Consider the feat of taking an earth-like
environment to the moon and back to support astronauts. That activity of environmental
creation emanating from human ego-activity and desires affects individual and societal
behaviour. This can result in excessive exposure to radiation or carcinogens from both the
natural (e.g. sunbathing) and directly man-made environments, including substances
ingested or inhaled. Here again is a way in which shortcomings in the higher activities of
the human constitution create the conditions for disease.
One criticism made by a statistician of the Scandinavian twin
study is that it was not possible to say if a particular case of cancer was entirely
environmental or genetic.25 This facile criticism can be levelled at all
genetic risk statistics. Already clinical geneticists can detect whether people carry in
their DNA certain precisely characterised gene sequences associated with disease. Finding
more of those sequences is an intended spin-off of the HGP. Screening for them will become
a fast routine automated procedure in the near future. Already DNA micro-arrays or chips,
coated with tens of thousands of different short DNA sequences called DNA probes can be
dipped in a solution of a patient's own DNA. If a sequence is present which is a
complementary match to a probe, it will stick to it through the tendency of the bases to
pair (see Fig. 1). A suitable fluorescent marker and laser detection system 'reads' which
disease-associated sequences are in the patient's DNA. But in many, if not most, instances
the result will enable the clinician to tell the patient only that they have a certain
statistical probability (or improbability) of getting symptoms of the associated disease.
This life's-a-lottery approach, will say very little if anything about the susceptibility
of the particular individual. Whereas pessimists will worry, perhaps becoming ill as a
consequence, optimists will simply ignore the information,26 indeed, perhaps
even have children in the face of a high risk of passing on the DNA sequence.27
However, even if this approach is looking in the wrong place for disease aetiology, the
general concept in the hoped-for fallout of the HGP that medical treatment will be totally
individualised cannot be dismissed out of hand. When medicine gradually realises that
illness is intimately connected with individual biography and is meaningful in such a
context, increasing individualisation of the study of the symptomology, even if only
initially at the DNA level, will be a complementary part of the quest for a treatment. But
the treatment, if DNA based, i.e. through gene therapy or pharmaceutical control of gene
expression, risks remaining within the current paradigm of treating merely symptoms, as
happened, for instance, when proponents of the germ theory of disease hit upon
antibiotics. Put simply, this bottom up approach needs to be complemented with the top
down approach for a truly holistic science and art of medicine to arise. But currently
almost all the wealth available for medical research goes on bottom up approaches like the
HGP.
Holtzman and Marteau, writing in the prestigious New England
Journal of Medicine draw on a large body of evidence from genetics to argue that
"the new genetics will not revolutionize the way common diseases are identified or
prevented. Mapping and sequencing the human genome will lead to the identification of more
genes causing Mendelian (see Box 1) disorders and to the diagnostic and predictive tests
for them.28 But most of the human bodily form and function cannot be accounted
for in terms of Mendelian inheritance, that is, the linking up of characteristics or
traits which geneticists call phenotypes, abstracted from the totality of the
organism, with associated inheritable markers called genotypes. Note that
consistent with the foregoing discussion we do not speak of genes for or causing
a disorder here, but of association. The gene, more correctly viewed, is one factor
in an organism's environment, albeit its intimate environment, which along with other
bodily substances, nutrients, warmth, light, its parents, society etc is essential for
that organism to manifest. Amongst the bodily substances the imprinting of the form is not
a monopoly of any specific substance. Of course, disturbing or accidental influences from
any of the factors, including genes can impede full manifestation of the ideal form or
type of the organism, or individuality in the case of humans. The association between a
particular gene and its related disease can be stronger or weaker as measured by the
statistical tests used in genetics. Mendelism works well for something like Huntington's
disease which is due to a single, highly penetrant dominant gene. By high
penetrance we mean here that if the gene is present, the observable characteristics
associated with it, in this case a disease pattern, are highly likely to manifest.
Furthermore, if the person affected has only one copy of the associated 'faulty' gene,
i.e. on only one chromosome of the pair, in order for the disease to manifest the gene
associated with it must be dominant. Holtzman and Marteau also challenge the notion that
the HGP ushers in the era of genomic medicine by pointing out that in over 40 years since
the molecular basis of the heritable disease sickle-cell anaemia was discovered, no
definitive treatment for it has emerged. They point out that many combinations of genes
strongly associated with disease, environmental factors and behaviour could all lead to
the same pathogenic effect. And we can concur with this from our view that disease or its
absence is administered at a much higher level in the organism's hierarchy than at the
level of 'external factors', amongst which we must include genes. And that higher level,
human individuality working alone or socially with others creates the differences in
lifestyle, environment and social structure to which Holtzman and Marteau attribute the
lion's share of the conditions for disease as compared with genetic differences. And as
regards our early years, it would not be stretching the imagination to regard our
surroundings and the people in it as part of our inheritance.
If neither genomic medicine nor knowing what it is to be human are
to be likely benefits to humanity from the HGP we can safely conclude that it would have
been better to spend the billions on developing a holistic approach to understanding
disease in parallel with educating to change lifestyles as well as reducing social
deprivation and environmental pollution. But this kind of investment in the future would
mean a widespread change in the mind set of a society that has largely accepted the DNA
myth and raised it to the level of a cultural icon.29 Such a change would take
many decades to effect, rather than the 10 years for the technical fix of the HGP. We must
now watch the project reach its conclusion before the limitations of its approach come
home to its proponents, as indeed we had to do with atomic power. Meanwhile, I'm with Tom
Shakespeare when he says 'William Shakespeare still has more to tell us about human nature
than genomics'.30
David Heaf is UK co-ordinator of Ifgene – the International
Forum for Genetic Engineering. (Ifgene home page)
Acknowledgements: The author wishes to thank his
wife Pat Cheney for her help with the text; Rudolf Saacke for his bibliographic search on
his electronic database of the Rudolf Steiner Gesamtausgabe and Don Powell
of the Sanger Centre, Cambridge for advice on the HGP.
Notes
1. New Scientist, 1 July 2000, p4. www.newscientist.com
2. Wellcome News, issued by the Wellcome Trust, Issue 20, 3rd Quarter,
1999, pp14-15.
3. Eric Lander, the Whitehead Institute, quoted in New Scientist, 1 July 2000,
p4..
4. New Scientist, 20 May 2000, p16
5. Darnovsky, M. & Hayes, R. (2000) Techno-Eugenics Email List newsletter,
No. 10, August 4. Subscription enquiries to teel@adax.com.
6. Wellcome News, issued by the Wellcome Trust, Issue 23, 2nd Quarter,
2000, pp2 & 8/9.
7. Mike Dexter, Wellcome Trust, quoted in New Scientist, 1 July 200, p4.
8. Annas, G. J. & Elias, S. Eds. (1992) Gene Mapping Using Law and Ethics as
Guides. Oxford University Press. (Enhanced proceedings of a workshop, January 1991,
Bethesda, Md.)
9. Appleyard, B. (1999) Brave new Worlds: Genetics and the Human Experience.
HarperCollins Publishers, London. Review
10. Steiner, R. (1886) A theory of knowledge implicit in Goethe's world
conception. Anthroposophic Press, New York. 1978
11. Steiner, R. & Wegman, I. (1925) Fundamentals of therapy. An extension to the
art of healing through spiritual knowledge. Rudolf Steiner Press, London, 1967.
12. Watson, J. D. (1989) quoted by Susan Lindee in The Future of DNA, Wirz, J.
& Lammerts van Bueren, Eds. Kluwer Academic Publishers. Chapter 3: The cultural powers
of the gene – identity, destiny and the social meaning of heredity. confer.htm
13. Steiner, R. (1910) Macrocosm and microcosm, 11 lectures, Vienna, 21-31 March,
lecture 5. Rudolf Steiner Press, London, 1968.
14. Lawson, D. (1995) All you need is life. The Spectator, 17th June.
15. Heaney, S. J. (1992) Aquinas and the presence of the human rational soul in
the early embryo. In Abortion: a new generation of Catholic responses. Chapter 3.
Heaney, S. J. Ed. Pope John XXIII Medical-Moral Research and Education Center, Braintree,
Mass.
16. Watt, H. (1998) The Origin of Persons. In Identity and Statute of Human Embryo.
Proceedings of the 3rd Assembly of the Pontifical Academy for Life, Vatican
City, 14-16 February 1997. Correra, J de D. V. & Sgreccia, Eds. Liberia Editrice
Vaticano.
17. Weihs, T. J. (1986) Embryogenesis in myth and science. Chapter 9. Floris Books,
Edinburgh. The spirit-soul elements called by Steiner 'ego', 'astral' and 'etheric',
correspond to Thomas Aquinas' concepts of rational, sentient and vegetative souls
respectively.
18. See for instance Vines, G. (1998) Hidden Inheritance. New Scientist, 28
November, pp27-30. www.newscientist.com
19. Watson, J. D. (1989) Ibid.
20. Van der Wal, Jaap (1997) Back to the future – towards a spiritual attitude for
managing DNA. In The Future of DNA, Wirz, J. & Lammerts van Bueren, Eds. Kluwer
Academic Publishers. Ch. 5. confer.htm
21. See for instance Popplebaum. H. (1977) New light on heredity and evolution,
St George Publications, Spring Valley. Popplebaum takes a phenomenological approach to
heredity which generally stands the test of modern developments in molecular genetics, but
the emphasis of some of his conclusions would need revising in the light of some more
recent discoveries.
22. Bott, V. (1978) Anthroposophical medicine. An extension of the art of healing.
Rudolf Steiner Press, London. The subtle intermediate bodies referred to are 'astral' and
'etheric' (see also note 17).
23. Popplebaum, H. Ibid.
24. New Scientist, 22 July 2000, p10 reporting on Lichtenstein, P. et al. (2000)
New England Journal of Medicine 343, July 13, 78-85.
25, Richard Peto, quoted in New Scientist (see note 24).
26. Bascou, J. (1999) Les progrès de la génétique: un défi pour notre temps.
Première partie: L'entrée dans l'ère du génie génétique. Association de Patient de
la Mèdecine d'Orientation Anthroposophique. Bulletin no. 38, Summer 1998, pp3-10. artsf.htm
27. Shakespeare, T. (2000) My two children and I share a genetic abnormality. So why
does this week's scientific milestone scare me so much? Daily Mail, 28 June, p12.
28. Holtzman, N. A. & Marteau, T. M. (2000) Sounding board: Will genetics
revolutionize medicine? The New England Journal of Medicine, 343(2), pp141-144.
29. Nelkin, D. & Lindee, M. S. (1995) The DNA Mystique. The gene as cultural icon.
Freeman, N.Y.
30. Shakespeare, T. (2000) http://www.sanger.ac.uk/HGP/draft2000/commentary.shtml
(Tom Shakespeare is the director of the Policy, Ethics and Life Sciences Research
Institute, Newcastle University and was born with a genetic abnormality called
achondroplasia – he is 4ft 5in high.)
© David Heaf, 14th October 2000
To Ifgene articles page
To Ifgene Home Page
Image files: heaf31.gif, heaf32.gif, heaf33.jpg, heaf34.jpg
The URL of this article is heaf3.htm
Other articles by this author can be found in the articles
section of Ifgene web site and at http://www.heaf.freeuk.com/articles.htm
.