LES PROGRES DE LA GENETIQUE : UN DEFI POUR NOTRE TEMPS
Deuxième Partie : L'histoire du concept de gèneJacqueline Bascou
Sommaire
L'histoire du concept de
gène
Le prologue
I Gregor
Mendel (1822-1884): fondateur de la méthode génétique
Des choix expérimentaux déterminants
Le langage de Gregor Mendel
A caractère constant, gène constant
II Thomas
Morgan (1866-1945), la génétique devient une science
Les gènes prennent place sur les
chromosomes
Les bons et les mauvais gènes
III Le
triomphe de la génétique moléculaire des années 1960-70
La surprise de l'ADN, substance des
gènes
Les merveilles de la double hélice
L’unité du vivant dans
le code génétique
IV La certitude et
le doute de la génétique actuelle
L’engouement pour la génétique
Les doutes du "tout
génétique"
Bibliographie
L'amélioration des espèces végétales ou animales semble avoir été pratiquée depuis des temps très anciens par les hommes. Il est vraisemblable que pour cela les plantes et les animaux ayant les meilleures performances aient été choisis pour obtenir la génération suivante. Probablement l'homme a guidé les fécondations quand il le pouvait. L'histoire du troupeau de moutons de Jacob dans l'Ancien Testament en est un exemple (Genèse 30). Jacob élevait les moutons de son beau-père Laban, ceux-ci étaient noirs ou tachetés. Il demanda à Laban de continuer à garder son troupeau et d'avoir comme salaire les moutons tachetés. "Toutes les fois que les brebis vigoureuses entraient en chaleur, Jacob plaçait les branches dans les auges sous les yeux des brebis, pour qu'elles entrassent en chaleur près des branches. Les brebis entraient en chaleur près des branches, et elles faisaient des petits rayés, tachetés et marquetés. Quand les brebis étaient chétives, il ne les plaçait point; de sorte que les chétives étaient pour Laban et les vigoureuses pour Jacob". C’est le désir d’avoir des brebis vigoureuses qui guide l’action de Jacob. Son action repose sur l'idée d'une hérédité de la vigueur de la mère à ses descendants. Cette manière de faire apparaît naïve de nos jours car l’hérédité n’est pas si simple. On peut penser qu’il a fallu beaucoup d’essais dus à des volontés d’hommes pour obtenir les variétés actuelles.
I Gregor Mendel (1822-1884): fondateur de la méthode génétique
Des choix expérimentaux déterminants
Gregor Mendel, fondateur de la génétique, rompt avec le discours flou ou poétique du passé. Il reçut une formation scientifique à l’Université de Vienne et devint supérieur du monastère Augustin de Brünn (aujourd’hui en Autriche). Là, il dirigea un jardin botanique où il mena ses célèbres expériences sur les pois.
L'observation dont il est parti devait être courante à son époque mais elle l’avait particulièrement frappé. Voilà l'introduction de son article. "C'est en procédant sur des plantes d'agrément, à des fécondations artificielles destinées à obtenir de nouveaux coloris, que l'on a été amené aux recherches qui vont être exposées ici. La régularité remarquable avec laquelle revenaient les mêmes formes hybrides, toutes les fois que la fécondation avait lieu entre les mêmes espèces, donna l'idée de nouvelles expériences dont le but serait de suivre les hybrides dans leur descendance." Gregor Mendel pressent dans cette régularité remarquable l’existence d’une loi de l’hybridation. Son originalité réside alors dans le choix des conditions pour conduire ses expériences. Ces conditions vont être déterminantes pour l’avenir de la génétique. Il les énonce ainsi : "Le choix du groupe végétal à utiliser dans les recherches de ce genre doit être fait avec les plus grandes précautions si l'on ne veut, dès le début, compromettre toute chance de succès. Les plantes d'expériences doivent absolument satisfaire à certaines conditions : elles doivent posséder des caractères différentiels constants; il faut que, pendant la floraison, leurs hybrides soient naturellement, ou puissent facilement être mis à l'abri de toute intervention de pollen étranger; les hybrides et leurs descendants ne doivent éprouver aucune altération notable de fertilité dans la suite des générations". Tous les organismes modèles choisis pour les études génétiques futures répondront à ces trois conditions, par exemple, la levure de boulangerie, la souris, la mouche du vinaigre (drosophile). Pour tous ces organismes, il est possible de diriger les croisements, d’avoir des descendances abondantes pour que l’on puisse utiliser les lois de probabilité et d’avoir des organismes ayant des traits de caractères alternatifs constants.
Parmi les 34 variétés de petit pois commun (Pisum sativum) qu’il cultivait, il en choisit qui présentaient des caractères alternatifs stables et les hybrida artificiellement entre elles. Par exemple, il hybrida des plants à pois jaunes et ronds avec du pollen de plants à pois verts et ridés. Puis il laissa les hybrides s’autoféconder naturellement pour obtenir une deuxième génération, et toujours par autofécondation il obtint une troisième et une quatrième génération. Tous les hybrides de première génération étaient jaunes et ridés. Il dira alors que jaune et ridé sont des caractères dominants et que vert et rond sont récessifs. De ce fait, il déduisit la loi de l’uniformité de la première génération (première loi de Mendel). A la deuxième génération, surprise, les deux traits de caractères parentaux réapparaissent à l’identique et dans des proportions régulières qui définissent une seconde loi, la loi de ségrégation. Avec beaucoup de ténacité, il continua l’analyse des plants de troisième et quatrième génération et il conclut à l’existence de facteurs héréditaires transmis par les cellules sexuelles. Ces facteurs héréditaires seront nommés gènes en 1909 par le botaniste danois Wilhelm Johannsen et les deux formes alternatives du gène seront nommés allèles. Enfin, dans les hybridations impliquant plusieurs caractères, chaque caractère semblant se combiner indépendamment de l’autre, il en déduisit la loi d’assortiment indépendant (troisième loi de Mendel). Ses travaux publiés en 1865 eurent une petite audience. Mais en 1902, les lois qu’il avait établies furent retrouvées et étendues aux plantes, aux animaux et à l’homme. Les lois de Mendel sont maintenant enseignées dans les écoles. Mais l’on oublie que ces lois ont été établies dans un cadre particulier. Ceci a des conséquences au moment où la génétique prend un essor considérable avec le génie génétique.
Gregor Mendel a pris grand soin de choisir des traits de caractères constants afin de pouvoir les nommer par des symboles. Il est clair que les caractères eux-mêmes ne l’intéressaient pas, ce sont des marqueurs d’hérédité qu’il a choisis. Dans l’exemple précédant, il nommera A les pois jaunes et a les pois verts et pour le caractère de la forme, il dira B pour pois ridés et b pour pois ronds (les grandes lettres étant attribuées aux caractères dominants). Ces appellations vont être transférées aux cellules sexuelles qui transmettent ces caractères. Ainsi la forme hybride formée par un ovule d’une plante A et du pollen d’une plante a sera nommée Aa bien qu'elle présente un caractère A (caractère dominant). C’est une manière d’indiquer que l’information du caractère a reste à l'état latent dans l'hybride puisqu’il réapparaît à la génération suivante. Finalement il transfert l’appellation A ou a à un couple de facteurs transmis par les cellules sexuelles. Chaque cellule sexuelle n’en ayant qu’un seul exemplaire, les plantes issues de fécondation en auront deux. Ainsi naît l'idée de facteurs constants (gènes) qui existent par paires (allèles) et qui déterminent les traits de caractère. Ces facteurs se séparent dans les cellules sexuelles et se réunissent lors des fécondations. On dira plus tard qu’il y a au cours d’un cycle de reproduction, alternance entre une phase haploïde (où chaque gène est en un exemplaire) et une phase diploïde (où chaque gène est en deux exemplaires). Le travail de Gregor Mendel est réellement fondateur puisque la majorité des concepts de la génétique formelle (indiqués en italique) est introduite.
A caractère constant, gène constant
Toutes les analyses génétiques futures se focaliseront sur des caractères constants car ils sont plus simples à étudier. Cette simplicité vient du fait qu'ils varient peu avec les conditions extérieures et que souvent un seul gène est concerné (déterminisme monogénique). Cette constance sera encore accentuée dans les études en laboratoire où tous les paramètres extérieurs sont contrôlés. De manière commode, le gène sera nommé par le caractère qui varie, par exemple, gène de la couleur ou de la forme de la graine. En fait, les gènes qui se prêtent facilement à la méthode scientifique représentent une classe bien particulière. Car pour tout observateur de la nature n’est-il pas évident qu’il y a plus de caractères inconstants que de caractères constants et que ce qui détermine la vie est complexe !
La relation entre le gène et le caractère n’a pas été soulevée par cette approche. Cependant l’on dira (et l’on dit encore) que le gène gouverne ou détermine le caractère , car quand il varie, le caractère varie. Ainsi, ce que les généticiens entendent par déterminer un caractère n’est pas ce que l’on entend habituellement dans le mot déterminer en français. Le concept de gène est cependant posé : les gènes seront considérés comme des entités autonomes qui déterminent les caractères et se trouvent dans les cellules sexuelles. Ils se transmettent sans mélange et ne sont influencés ni par l’hybridation ni par le milieu. On peut se demander si l’autonomie (la constance) attribuée aux gènes n’est pas une conséquence du choix d’étudier des caractères constants.
II Thomas Morgan (1866-1945), la génétique devient une science
Les gènes prennent place sur les chromosomes
Thomas Morgan, biologiste américain, inaugura les débuts de la génétique en tant que domaine scientifique propre. Il choisit la drosophile, dite communément mouche du vinaigre qui répond aux trois conditions de Gregor Mendel énoncées ci-dessus. Les travaux de Thomas Morgan mirent en évidence que si certains couples d'allèles se combinaient indépendamment dans la descendance d'hybrides, comme Gregor Mendel l'avait observé (troisième loi de Mendel), d'autres présentaient une tendance à rester associés ensemble. En prenant en compte le degré de liaison, il devenait possible d'aligner les gènes et de les rassembler en groupes de liaison dont le nombre est défini pour chaque espèce. L'idée que les groupes de liaison correspondaient aux chromosomes du noyau des cellules fut proposée sur la base de la convergence des comportements des gènes et des chromosomes lors de la reproduction sexuée. Thomas Morgan reçut un prix Nobel en 1933 pour la théorie chromosomique de l'hérédité. Chaque gène pouvait se définir par une place sur un chromosome (appelé locus). Leur représentation se précisait et l’hérédité devenait chromosomique.
Un changement important du langage eut lieu à cette époque, mais il passa inaperçu. Depuis Gregor Mendel, les traits de caractères alternatifs étudiés étaient trouvés dans la nature. A partir des années 30, Thomas Morgan et les généticiens commencèrent à utiliser des mutagènes, en particulier les rayons X, pour faire varier les caractères, les variants obtenus étant des mutants. Les altérations qui auront touché les gènes des cellules sexuelles seront transmises à la descendance, on les nommera mutations. Pour les drosophiles, Thomas Morgan procéda ainsi : des mouches de sexe mâle, toutes semblables, issues d’une lignée de référence dite souche sauvage, furent traitées aux rayons X. Ces mouches traitées furent croisées avec des femelles de la souche de référence et leurs descendances observées sur plusieurs générations. A chaque génération, les descendants qui présentent un trait de caractère modifié seront recherchés. Par exemple, ils auront des yeux blancs au lieu d’avoir des yeux rouges comme dans la souche de référence. Si la modification est héréditaire, on dira qu’une mutation est apparue lors de la mutagénèse. La mutation portée par ces mouches mutantes révèle l'existence d'un gène dont la localisation et la fonction seront ensuite recherchées. La mutation étant un marqueur d’un gène, il sera plus commode de nommer le gène concerné par le trait de caractère mutant. Par exemple, le gène muté chez les mouches à yeux blancs sera nommé gène white (blanc). Gregor Mendel aurait appelé ce gène, gène de la couleur de l’œil. Ce changement dans la manière de nommer un gène s'est avéré commode mais aussi nécessaire, car de nombreuses mutations affectant la couleur de l’œil furent trouvées en des endroits (locus) différents des chromosomes, donc dans des gènes différents. Il était ainsi montré que de nombreux gènes peuvent déterminer un même caractère. Il est important de comprendre que cette manière de nommer les gènes peut conduire à un contresens quant à la compréhension de la fonction des gènes. Les mutations provoquées par des mutagènes étant des altérations des gènes qui conduisent souvent à leurs inactivations, cela revient à nommer le gène par ce qui apparaît quand le gène n’est pas fonctionnel. C'est comme si on nommait le cœur, l'organe de la mort puisque quand il s'arrête de fonctionner, on meurt !
Cette dérive du langage avait déjà commencé avec l'étude de maladies héréditaires chez l'homme. Par exemple, on commençait à parler du gène de l'hémophilie. Ce langage continue d’être utilisé de nos jours surtout dans la presse. Ceci a pour conséquence de donner une moralité aux gènes. Il y a les bons gènes qui par mutation ne vont pas donner de maladie ni entraîner la mort et il y a les mauvais gènes qui peuvent donner la maladie ou la mort. L'idée de thérapie génique naît ainsi, il faut réparer les gènes malades. Le danger de l’eugénisme des années 30 réapparaît sous une autre forme qui pourrait paraître "moralement correcte".
III Le triomphe de la génétique moléculaire des années 1960-70
La surprise de l'ADN, substance des gènes
La question de la substance des gènes fut abordée par des expériences de transformation chez les bactéries réalisées en 1928 par l'anglais Griffith. Il mit en évidence que des bactéries mortes pouvaient transmettre des caractères à des bactéries vivantes et que cette transformation devenait héréditaire. La nature de la substance transformante fut posée. Les différents constituants des bactéries étant connus, ils furent testés les uns après les autres. Et c’est avec surprise que l’une des substances contenue dans les chromosomes, l’ADN, soit identifiée comme le facteur transformant en 1944 par O. T. Avery, C. M. McLeod et M. J. McCarty. Pourquoi cette surprise ? Cette substance des chromosomes était connue des biologistes et des chimistes, mais elle avait été jugée trop simple pour contenir une information génétique. L'ADN purifié apparaît en effet comme une substance amorphe, solide, incolore, filamenteuse que l'on peut rouler comme des spaghettis sur une baguette. En masse, elle est parfois appelée la pieuvre. L’analyse chimique avait révélé un polymère formé d’un petit nombre de composants. Comment une substance aussi simple pourrait-elle porter l’information du monde vivant ? Ce devait être l’énigme des recherches ultérieures.
Les merveilles de la double hélice
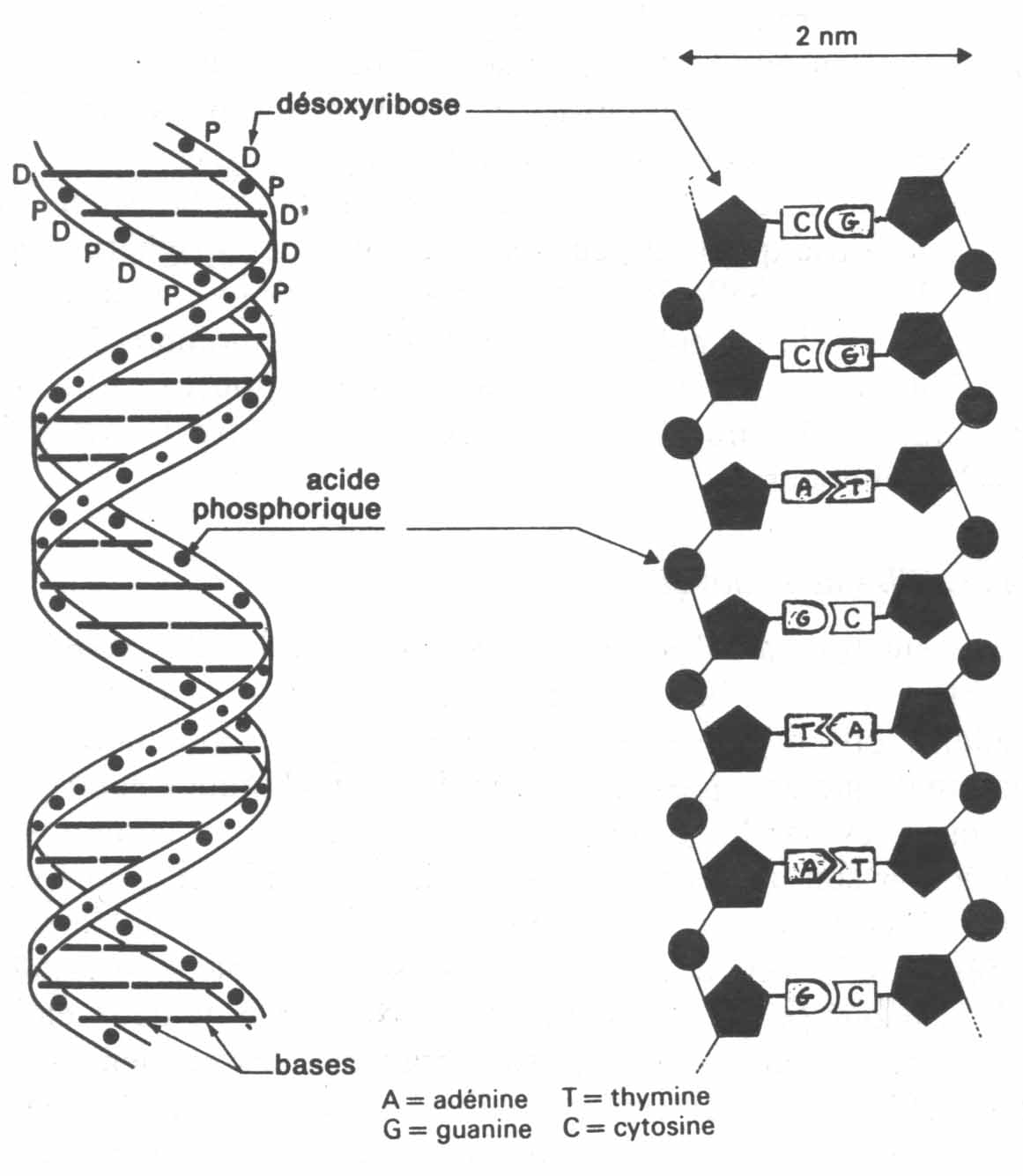
La représentation que l’on se fait aujourd’hui de l'ADN, celle dite de la double hélice, est beaucoup plus complexe mais à l’inverse de la précédente elle présente la beauté d’un cristal répondant ainsi au besoin mythique de représenter la vie. Le nom chimique étant compliqué (acide désoxyribonucléique), cette substance est communément nommée par son abréviation ADN. Elle est représentée comme une double hélice très régulière formée de deux brins complémentaires appariés de telle sorte que le modèle utilisé ressemble à un escalier en spirale (voir la figure). Chaque brin est constitué d’un axe formé de deux éléments (sucre et phosphate) sur lequel sont branchés des éléments qui eux sont variables. Ces éléments variables sont au nombre de quatre, ce sont les bases nucléiques. Ces quatre bases sont nommés par leurs abréviations chimiques : A pour adénine, T pour thymine, G pour guanine et C pour cytosine. L’adénine et la guanine appartiennent à une même famille chimique, alors que la thymine et la cytosine appartiennent à une autre famille. C’est par ces bases que les deux brins sont appariés car elles sont complémentaires deux à deux (ce sont les bases appariées qui forment les marches de l’escalier). Dans les deux brins, en face d'un A il y a toujours un T, et en face d'un G il y a toujours un C. Ainsi, on définit une molécule d’ADN par la séquence de ces éléments variables. Par exemple, l'un des brins peut avoir en un endroit la séquence CTGCCGATAAAG. Par la loi de complémentarité, l’autre brin sera GACGGCTATTTC. James Watson, Francis Crick et Maurice Wilking reçurent le prix Nobel en 1962 pour le modèle de la double hélice de l'ADN.
Schématisation de la molécule d'ADN et d'un ADN supposé déroulé
L’unité du vivant dans le code génétique
Comment imaginer que l'information génétique du monde vivant soit écrite dans un alphabet à quatre lettres (A, T, G, C) appartenant à deux familles chimiques ? Ce serait comme un langage écrit dans un alphabet à deux voyelles et deux consonnes ! Ce sera seulement en 1961 qu’il sera établi que les lettres de l'ADN se lisent trois par trois (triplet), ce qui donne un alphabet à soixante quatre lettres. La traduction de ce message dans les cellules sera résolue en même temps et conduira à ce que l’on appelle aujourd'hui le code génétique. Il sera montré que l’ADN est traduit, dans la majorité des cas, en protéines par l’intermédiaire d’un acide nucléique messager. Les protéines, elles, sont des molécules écrites dans un alphabet à vingt lettres qui sont les vingt acides aminés. Le code génétique sera le dictionnaire donnant la correspondance entre les soixante quatre triplets de base et les vingt acides aminés. En reprenant l'exemple ci-dessus, la séquence d’ADN : GAC GGC TAT TTC se traduira en une séquence d'acides aminés : leucine, proline, isoleucine, lysine. Un enthousiasme triomphant viendra du fait que, avec quelques arrangements, ce code génétique semble s’appliquer à tous les organismes. On dira que le code génétique est universel. Ce qui fit dire à Jacques Monod (prix Nobel en 1965) : "ce qui est vrai pour les bactéries est vrai pour l'éléphant". Le mendélisme et le darwinisme trouvèrent là un support convenable. La théorie unificatrice du néodarwinisme fut établie. Elle tient compte à la fois des lois de Mendel et du rôle de la sélection naturelle dans l’évolution (darwinisme), les objets sur lequel s’exercent ces lois ou la sélection étant les gènes constitués d’ADN. Un large consensus s’est formé autour de cette théorie et s’y opposer relève d’un combat d’arrière-garde. Dans cette théorie, le gène est, généralement, une séquence d’ADN portant l’information pour une protéine. Les caractères observés ont encore plus disparus de la conscience des chercheurs, tout est pensé maintenant sur un plan moléculaire.
IV La certitude et le doute de la génétique actuelle
L’engouement pour la génétique
Un monde génétique virtuel s'est créé. L'ADN est vu comme une substance dans laquelle on peut lire le passé (l'évolution du vivant), le présent (compréhension des phénomènes ou diagnostic génétique) et l'avenir (prédiction des désordres qui pourraient survenir). Le génome étant le livre de la vie, il suffit maintenant de le lire. Ainsi tous les efforts sont tournés vers le séquençage des génomes et les premiers organismes génétiquement modifiés (OGM) sortent des laboratoires.
Dans les universités, un nouvel enseignement est mis en place : "la génomique". Dans cette nouvelle science, un vocabulaire est utilisé qui témoigne de nouvelles interrogations sur le sens de ce qui apparaît au niveau ADN. Par exemple, on parle d’ADN inutile (si on ne sait à quoi il sert), orphelin (si on ne trouve pas d’homologue chez d’autres organismes), égoïste (idée développée par Richard Dawkins qui donne aux gènes valeur d’individualités œuvrant pour leurs propres comptes ; les organismes deviennent alors les vecteurs des gènes !), etc …
Cet engouement pour la génétique vient de plusieurs pôles très différents, en voici trois. Le premier est banal, il repose sur le besoin des hommes d’un monde meilleur. Qui ne souhaite pas un monde sans maladie et produisant de bonnes choses en quantité ! Depuis Charles Darwin, la nature est considérée comme le fruit d’une évolution. L’existence de ce que l’homme voit comme des imperfections conduit à l’idée que l’évolution est le produit du hasard et de la nécessité (Jacques Monod, 1970), que c’est du bricolage (Fançois Jacob, 1977) dont le produit peut être "mal fichu". Le désir d’améliorer le "mal fichu" sert à justifier les manipulations génétiques et donne de la noblesse aux intentions poursuivies.
Le deuxième agrément vient de la recherche scientifique et de la confiance qu’on lui accorde. En maintenant le cadre évoqué ci-dessus, des succès surprenants ont été obtenus largement annoncés dans la presse. Il est clair que trouver ce que l'on cherche, même si c’est très rare, apparaît comme une garantie de la validité de la pensée qui l'a produite : "ça marche" donc c'est vrai ! C’est l’engouement du "ça marche !". La rareté de ce qui est produit pourrait laisser entendre que ces résultats sont contraires à la nature, pourtant ils existent, la nature peut les produire. Il est de plus en plus montré que des transferts génétiques dit horizontaux (par opposition aux transferts verticaux de parents à descendants) existent naturellement par le biais des micro-organismes. Ceci soutient l’idée que les biotechnologies génétiques font plus efficacement ce que la nature sait faire.
Le troisième pôle favorable au génie génétique est celui du monde économique. Les sociétés multinationales s'en sont emparées et ont investi d'énormes moyens qui sont à la mesure de l'espérance des bénéfices futurs. Il suffit de lire leurs messages publicitaires pour comprendre que ces sociétés font tout pour défendre l'idée que les manipulations génétiques sont en accord avec la nature et qu’elles vont nous conduire à un monde meilleur. Ce pouvoir là est prêt à entretenir toute illusion qui l’arrange. Le "tout génétique" permet l’instauration d’un nouveau pouvoir, celui du gène. Il permettra l’appropriation des fonctions biologiques, par brevets.
Les doutes du "tout génétique"
L'illusion d’une vision simpliste dite du "tout génétique" est maintenant dénoncée aussi bien par les scientifiques que par les citoyens mais pas par les sociétés privées de biotechnologies qui sont impliquées économiquement. Les études des vingt dernières années qui portent davantage sur les plantes, les animaux et l'homme que sur des micro-organismes ont apporté de nouveaux résultants. La notion de gène a dû être révisée. Car on s’est aperçu que les gènes pouvaient être morcelés, qu’il était difficile de leur attribuer des frontières, qu’ils étaient parfois incomplets et pouvaient être corrigés avant d’être traduits en protéines, qu’à une même séquence d’ADN pouvaient correspondre plusieurs protéines. Bref, malgré quelques tentatives, le gène ne pouvait plus se définir simplement par une séquence définie d’ADN portant une seule information. Le gène devenait un archétype susceptible de prendre des formes multiples. Le concept de gène sort ainsi d’une représentation matérialiste pure de même que celui de l'électron en physique.
L’idée que le génie génétique est une technologie banale est aussi remise en cause. Il devient clair que le génie génétique inscrit dans les organismes vivants des projets humains. Même si des mécanismes de transfert génétique horizontaux sont observés dans la nature, ce qui est fait par les manipulations génétiques va bien au-delà. Non seulement on passe les barrières d’espèces, mais on passe aussi la frontière des règnes du monde vivant (micro-organismes, végétaux, animaux, homme). De plus, les gènes greffés sont eux-mêmes des chimères. Ce sont des assemblages de séquences d’ADN provenant de divers organismes tels que le gène étranger puisse s’exprimer dans le contexte génétique de l’organisme hôte. Ainsi peuvent être obtenus des plants de tabac capables de produire de l’hémoglobine humaine. Presque tout est possible puisque le monde vivant considéré au niveau ADN est quasiment unique.
Une autre source importante du doute vient de la validité de la modélisation génétique. N’a-t-on rien oublié ? Le modèle est-il suffisamment saturé de réalité pour qu’il puisse être utilisé en agriculture et en médecine ? L’exposé ci-dessus montre qu’au cours de l’histoire de la génétique, la réalité sensible a, au contraire, été progressivement évacuée au profit d’une modélisation moléculaire. Le fait que ce qui paraît peut dépendre d’un contexte de vie a été complètement oublié. Il est souvent admis qu’à un génotype correspond un seul type d’organisme. L’oubli d’autres dimensions de la vie est naturellement le plus nettement ressenti quand il s’agit de l’homme et de soi en particulier. Est-ce que mon identité (ou mon avenir) est inscrit dans mes gènes ? Ce thème est largement développé par Craig Holdrege dans son livre : "Genetics and the Manipulation of Life, the forgotten factor of context". Mais sommes-nous prêts à redonner vie au contexte ? Aimons-nous la vie à ce point ? La vie dans sa complexité et sa mouvance nous insécurise, elle apparaît souvent épouvantable, voire dégoûtante. Il faut bien le reconnaître, notre pensée préfère le monde minéral, plus sûr. Aussi, il devient urgent de reconsidérer ce qu’est la vie et ce que nous voulons changer de celle-ci. Il faudra de l’audace et du courage pour sortir de cette caricature de la vie que la génétique nous présente actuellement.
- L’œuvre de Gregor Mendel. Extrait du Bulletin de «l’Union des Naturalistes de l’Enseignement Public », n° 3, Juillet-septembre 1961.
- Les Avatars du Gène, la théorie néodarwinienne de l’évolution de Pierre Henri Gouyon, Jean Pierre Henry et Jacques Arnoult. Regards sur la Science, Belin, 1997.
- Le Hasard et la Nécessité. Jacques Monod. Paris, Seuil, 1970.
- Evolution and Tinkering. François Jacob. Science, vol. 196, p 1161-1166 (1977).
- Le gène égoïste. Richard Dawkins. Paris, Armand Colin (Sciences), 1990.
- Genetics and the Manipulation of Life, the forgotten factor of context de Craig Holdrege. Lindisfarm Press, New York, 1996 (disponible au secrétariat d’Ifgene France, Colette Pradelle, La Commanderie, Amance, 10140, France)
La troisième partie : L’ADN et la Vie
Jacqueline Bascou: Jacqueline.Girard@ibpc.fr
Ces trois études ont été publiées dans la revue de l'APMA (Association des Patients de la Médecine d'orientation Anthroposophique), n° : 38, 39-40 et 41-42. Leur contenu a été presenté lors de l'Assemblée Générale de l'association APMA en 1997 par l'auteur. Ces revues peuvent être commandées au secrétariat de l'association à Colette Pradelle : La Commanderie, 10140, Amance, France.